La plante domestiquée



La plante domestiquée
Les différentes techniques de domestication de la plante
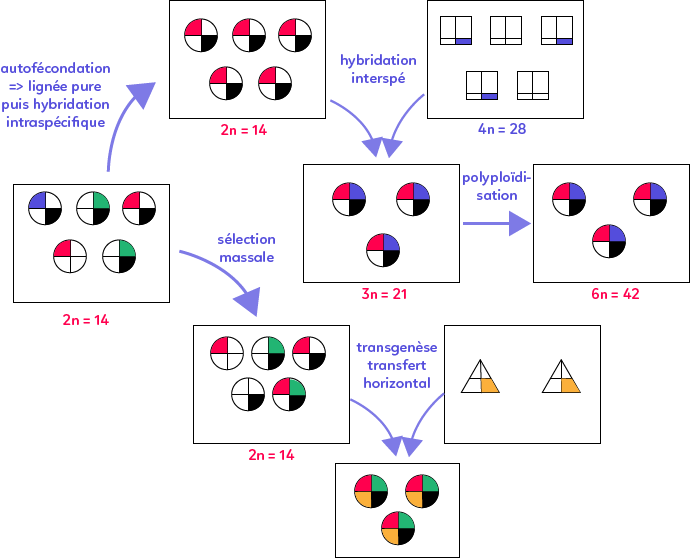
Une première technique d’amélioration des plantes consiste à pratiquer l’autofécondation, c’est-à-dire la reproduction d’un individu avec lui même, il n’y a ainsi pas de croisement. Une telle technique permet l’apparition d’une lignée pure, soit des individus homozygotes pour les gènes considérés. Une fois ces individus obtenus, on réalise un croisement aussi appelé hybridation intraspécifique, qui conduit à l’émergence d’individus identiques pour les caractères considérés. Ainsi, il est beaucoup plus aisé de cultiver de telles plantes car elles sont toutes identiques et présentent les caractères intéressants d’un point de vue agronomique.
Il est aussi possible de forcer l’hybridation entre deux espèces présentant des caractères d’intérêts différents, c’est l’hybridation interspécifique. On obtient alors une espèce stérile qui peut être rendue fertile grâce à la polyploïdisation et qui cumule les caractères des deux espèces.
On peut également choisir au sein de la population les individus apportant l’intérêt agronomique le plus grand : c’est la sélection massale. A l’issu de ce procédé, on obtient une population dont la diversité allélique a été réduite.
Enfin, une autre technique est la transgénèse : il s’agit d’un transfert horizontal de gènes. Une espèce présente un caractère intéressant codé par un gène que l’on isole puis que l’on insère dans le génome d’une autre espèce cultivée par transfection. On a alors une nouvelle espèce, dite organisme génétiquement modifié ou espèce transgénique, qui cumule ses anciens caractères mais surtout le caractère d’une autre espèce.
La sélection à des fins agronomiques
I. Exemple de la domestication du blé
Le blé est la principale céréale cultivée en termes d’hectares à l’échelle de la planète. Depuis que l’Homme se sédentarise, le blé est cultivé. Cela représente 800 000 ans de culture et de domestication des cultures donc avant même l’existence d’Homo sapiens (il y a 200 000 ans). Cette culture de céréales est donc extrêmement ancienne et est en lien avec la sédentarisation.

Quand on cultive des plantes, on réalise une sélection, non pas naturelle, mais artificielle. C’est une sélection massale qui est une action directe sur le phénotype des organismes. On cultive un champ de blé, on récolte des graines qu’on utilise dans l’alimentaire mais, sur toutes ces graines récoltées, on va en conserver certaines pour les mettre à germer pour l’année d’après. D’une année sur l’autre, on sélectionne les individus au sein d’une masse. On sélectionne les individus qui apportent les intérêts agronomiques les plus importants, alimentaires ou autres : ils produisent beaucoup, résistent à certains virus, au vent, etc.
Il existe deux types de blé (que l’on consomme) : le blé tendre (panification, farine) et le blé dur (semoule).
II. Comment a-t-on abouti au blé tendre ?
Le blé tendre, appelé Triticum aestivum comporte 42 chromosomes au total. Si on retrace l’histoire de ce blé, on estime qu’il y a à peu près 10 000 ans, ce blé est né de l’hybridation entre deux types de grains proches : l’Engrain à 2n = 14 et l’Égilope à 2n = 14.
Ces deux espèces différentes s’hybrident et donnent donc un hybride à « 2n = 14 ». En effet, grâce à la méiose, lorsque l’on est diploïde avec 14 chromosomes, on aura des gamètes haploïdes à 7 chromosomes. Lorsqu’on a des gamètes d’Engrain à 7 chromosomes et des gamètes d’Égilope à 7 chromosomes, on obtient des hybrides à 14 chromosomes. Ces hybrides sont stériles.
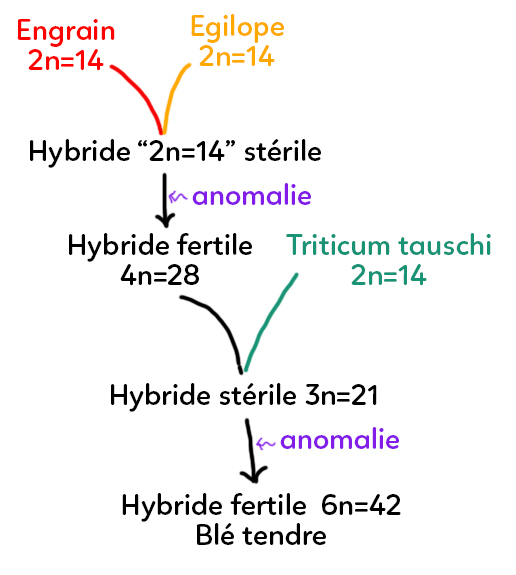
Par le biais d’une anomalie au moment de la cytodiérèse par exemple, on peut supposer que lors d’une mitose, les deux chromosomes se disjoignent mais la cellule ne donne pas deux nouvelles cellules filles. On obtient donc un doublement du stock chromosomique : on parle de polyploïdisation.
Cet hybride qui était stérile devient donc fertile suite à cette anomalie avec 4n = 28 (tétraploïdes). Cet hybride va donc ensuite se croiser avec une autre espèce de graminées : le Triticum tauschi, à 2n = 14.
Pour rappel de la méiose et de la fécondation, si on a une plante à 4n = 28, on va produire des gamètes diploïdes à 2n = 14. Le Triticum tauschi à 2n = 14 va donc produire des gamètes à n = 7. Cela donnera alors un hybride stérile à 3n = 21 chromosomes.
Cet hybride stérile ne peut donc pas avoir de descendance. Le hasard fait que l’absence de disjonction mitotique va donner un hybride à 6n = 42, fertile : c’est le blé tendre. Cet exemple d’hybridation entre espèces et de polyploïdisation est une illustration naturelle d’exemple d’hybridation entre espèces qui amène à l’espèce de blé tendre consommé par l’espèce humaine.
Finalement, la sélection à des fins agronomiques peut se faire de façon phénotypique (la sélection massale) et de façon génotypique ou génomique :
– Par hybridation naturelle ou artificielle : les ingénieurs agronomes forcent des hybridations entre espèces plus ou moins proches pour obtenir une nouvelle espèce qui va accumuler les avantages des deux espèces initiales.
– Par la polyploïdisation qui permet de rétablir la fertilité de l’hybride mais pas seulement. Si on prend les bananes qui sont polyploïdes, l’avantage est que cela augmente toujours le nombre de chromosomes et ainsi une cellule avec beaucoup de chromosomes sera une cellule plus grosse et à grande échelle on aura des fruits plus gros.
– On peut aussi parler des OGM (Organismes Génétiquement Modifiés) qui permettent d’insérer un gène d’intérêt d’une espèce donnée dans une autre espèce. Les exemples les plus courants d’OGM sont le maïs Bt et la tomate McGregor (plus commercialisée) qui proposait l’avantage de mûrir moins vite.