Microscope et théorie cellulaire



Microscope et théorie cellulaire
Microscope et théorie cellulaire
La notion de théorie cellulaire (le fait que les organismes vivants soient constituées de cellules) est une notion qui s’est construite au cours du temps. Sa construction a commencé au XVIIe siècle avec l’arrivée premiers microscopes.
I. Invention du microscope optique et première observation de cellules
Il n’y a pas de consensus sur la date et la personne qui a inventé le premier microscope. Pour information Jansen serait un des concepteurs des premiers microscopes optiques (MO). Grâce à un jeu de lentilles convergentes, l’idée était de faire converger des photons sur un porte-objet sur lequel un objet était observé en le grossissant. On date l’invention du microscope optique à la fin du XVIe siècle (1595).
Le mot « cellule » est associé à Robert Hooke. En 1665, il a placé du liège sous un microscope et observé des petites structures semblables à des boîtes les unes à la suite des autres. Il a utilisé le terme « cellule », du latin cellula, la cellule des moines. Cela signifie qu’on a initialement considéré qu’une cellule était un espace fermé, avec très peu d’échanges avec l’extérieur. Or le vivant est défini par sa capacité de convertir des énergies, d’être traversé par des flux de matière, d’énergie, d’informations. Il y a donc nécessairement un biais dans cette représentation initiale de la cellule : certes, elle est délimitée par une membrane, mais elle va laisser passer un certain nombre d’informations, elle n’est pas fermée. Cette première observation de cellules au microscope optique participe cependant à l’un des grands axes de cette théorie cellulaire : Hooke commence à dire que les êtres vivants sont composés d’une ou plusieurs cellules.
II. La cellule comme unité structurale du vivant
Au XIXe siècle, Schwann travaille sur des cellules de chorde (tissu embryonnaire qui donne notamment le système nerveux), donc sur de l’animal, et communique beaucoup avec un botaniste, Schleiden, qui utilise également un microscope dans ses travaux pour observer des végétaux. Tous deux convergent vers une idée : une cellule présente une unité dans sa structure, elle possède un noyau.
À l’élément de la théorie cellulaire stipulant que tout être vivant est constitué de cellules s’ajoute celui que la cellule est l’unité structurale du vivant, qui compose tout être vivant ; aussi bien les végétaux que les animaux. Il s’agit d’un tournant important dans l’élaboration de cette théorie cellulaire.
III. La théorie démentie de génération spontanée
Les travaux de Pasteur reprennent une idée de Schwann et étudient la notion de générations spontanées. Pendant longtemps, on pensait que les mouches qui arrivaient sur un cadavre en putréfaction naissaient spontanément. Pasteur teste par une expérience célèbre avec un col de cygne l’existence ou non de générations spontanées en chauffant un milieu riche en microorganismes et en empêchant l’air d’amener de nouveaux microorganismes dans le milieu. Par un jeu de témoins, il va prouver que l’air contient des microorganismes, qu’il n’y a pas de générations spontanées. Si un asticot se développe à un moment donné sur une viande, il n’est pas né de n’importe quoi : cela signifie qu’à un moment une mouche s’y est posée et a pondu des œufs.
C’est de nouveau un tournant important dans l’élaboration de la théorie cellulaire : c’est la fin de la croyance en la génération spontanée, toutes les cellules proviennent nécessairement de cellules préexistantes.
IV. L’avènement du microscope électronique
Au XXe siècle, dans les années 1930, un microscope plus puissant, au pouvoir de résolution plus important, est mis au point. Alors qu’avec un microscope optique on grossit jusqu’à 2 000 fois un objet, le microscope électronique à balayage peut grossir jusqu’à 100 000 fois un objet, et celui à transmission peut aller encore plus loin et grossir un objet jusqu’à plusieurs millions de fois. Il fonctionne par utilisation d’un faisceau d’électrons qui traverse l’objet, et n’est utilisable par conséquent que sur du tissu mort. Ernst Ruska en est l’un des concepteurs en 1932.
Le microscope électronique à balayage permet de visualiser la structure 3D de l’objet comme on peut le voir ci-dessous sur cette électronographie de cellule : on distingue le noyau par exemple.
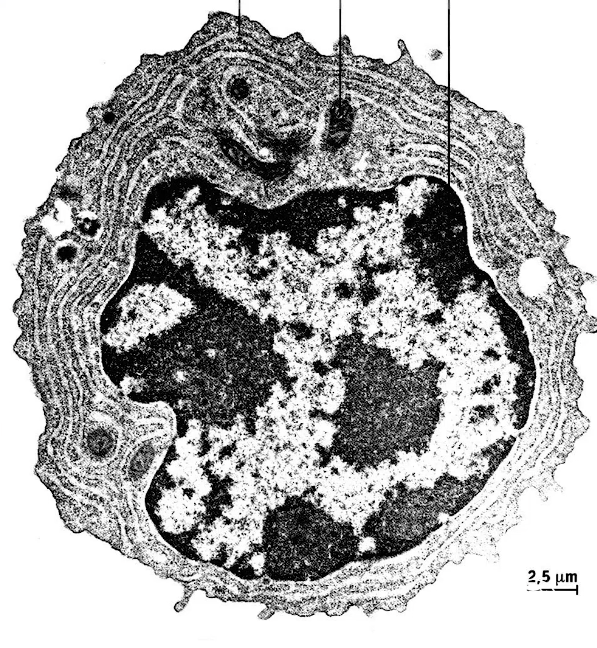
Le microscope électronique à transmission permet, lui, de distinguer jusqu’à la membrane de la cellule comme on peut le voir sur l’électronographie ci-dessous.
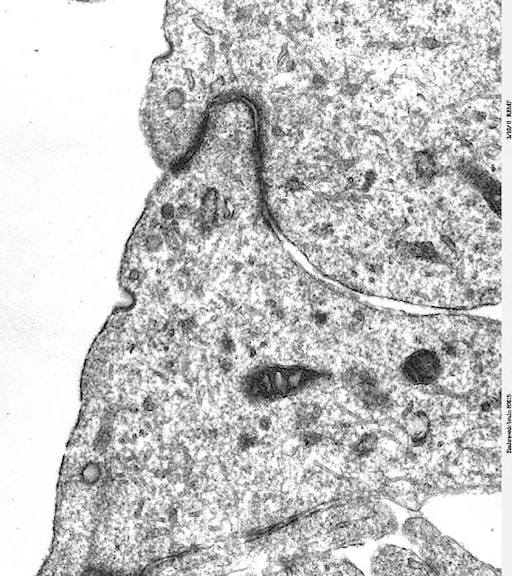
Ce niveau de résolution permettra notamment de comprendre l’ultrastructure de cette membrane, et de rompre définitivement avec l’idée de cellule du moine, de cellule fermée. Sa membrane est un élément semi-perméable qui va pouvoir laisser passer un certain nombre de substances, permettre aux cellules de communiquer avec les autres, qu’elles soient au sein d’un organisme pluri ou unicellulaire. Car une définition de la vie est d’être capable d’échanger. L’avénement du microscope électronique est donc un moment important dans la mise au point de la théorie cellulaire.
En 1972, grâce à la microscopie électronique et aux expériences de cryofracture et cryodécapage, Singer et Nicholson vont proposer le modèle de la mosaïque fluide de la membrane plasmique, dans l’idée que la cellule est délimitée par une membrane semi-perméable.
V. La notion de LUCA
Dans l’avènement de cette théorie cellulaire pour soutenir l’idée qu’une cellule provient nécessairement de cellules préexistantes, on peut citer les travaux de Forterre. En 1996, il énonce la notion de Last Universal Common Ancestor, l’ancêtre commun universel de tous les organismes vivants, qui est fait nécessairement d’une cellule, avec une membrane cytoplasmique semi-perméable, et un génome (de l’ADN ou de l’ARN) contenu à l’intérieur de cet organisme, puisqu’une des particularités de la vie c’est d’être capable de se reproduire. Donc toutes les cellules proviennent de cellules préexistantes qui se sont multipliées.
Organisation moléculaire de la membrane plasmique
Organisation moléculaire de la membrane plasmique
Une expérience connue est d’extraire de l’ADN d’oignon. L’idée c’est de broyer, donc d’exercer un action mécanique pour éclater les cellules et détruire un peu les membranes cellulaires. Souvent quand on fait cette expérience, on ajoute un liquide vaisselle ou un détergent. Ce liquide a pour fonction de dissoudre les lipides. Les membranes des cellules son constituées de lipides. C’est partiellement vrai. Nous allons voir la composition d’une membrane plasmique pour aboutir au schéma bilan ci-dessous.
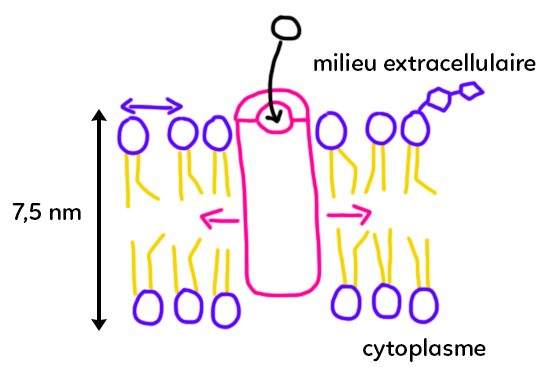
Ce schéma est compliqué et exhaustif, vous n’avez pas à connaître tout le vocabulaire.
I. Histoire : une bicouche et non une micelle de lipides
On pensait historiquement que la membrane était faite de lipides : mais de combien de couches sachant que ces lipides sont des glycérophospholipides (représentés ci-dessous) ?
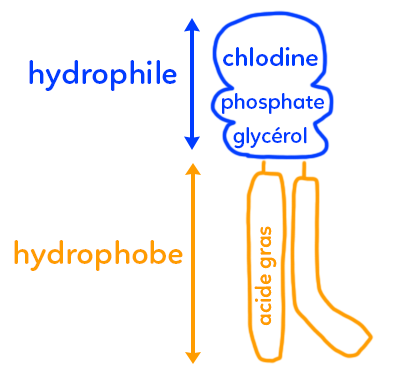
Nous sommes faits à 70 % d’eau. Nos cellules sont faites d’eau. Or une membrane, c’est lipidique et les lipides ne sont pas hydrophiles mais hydrophobes. Cela signifie que les lipides constitutifs des membranes présentent une partie hydrophile (qui aime l’eau), et une partie hydrophobe. La question est de savoir si les membranes cellulaires ne sont que des micelles de lipides (une seule couche de lipides) ou s’ils s’organisent en bicouche.
Dans les années 1920, on a extrait les lipides d’un globule rouge et on les a déposés sur une surface. On a constaté que lorsqu’on déposait tous les lipides membranaires d’un globule rouge on obtenait deux fois la surface du globule rouge. Cela signifie que la membrane d’un globule rouge est faite, non pas d’une, mais de deux couches accolées de lipides.
II. Structure du lipide membranaire
On part donc sur un modèle de bicouche lipidique, avec un lipide, ici un glycérophospholipide (hors programme). Ce lipide est composé d’une partie hydrophobe dite apolaire, riche en acides gras et d’une petite zone non négligeable, exposée vers le milieu extracellulaire qui est fait d’eau, mais aussi vers le milieu intracellulaire qui est lui aussi fait d’eau (selon que le lipide est positionné sur l’une ou l’autre des deux couches de la bicouche lipidique) : la zone hydrophile faite de choline (par exemple pour la phosphatidylcholine), d’un groupement phosphate et d’un glycérol.
On a ainsi deux couches de glycérophospholipides ou phospholipides : une couche qui présente sa partie hydrophile vers le milieu extracellulaire et une couche qui présente sa partie hydrophile vers le milieu intracellulaire (cytoplasme). Les parties hydrophobes se font face ce qui donne cette bicouche lipidique.
III. Les protéines membranaires
Mais ce n’est pas si simple : la membrane plasmique n’est pas constituée que de lipides. Comment a-t-on compris cela ?
Dans les années 1920-1930, Davson et Danielli travaillent sur la tension superficielle : la force qui donne à la surface d’un liquide un comportement élastique. Si la membrane plasmique n’est faite que de lipides, on va avoir une valeur théorique de la tension superficielle. Davson et Danielli mesurent la valeur de la tension superficielle d’une vraie membrane et observent une valeur inférieure à celle attendue sous l’hypothèse que la membrane plasmique n’est constituée que de lipides. Conclusion : la membrane plasmique n’est pas faite que de lipides.
Ils se disent alors que d’autres molécules que les lipides seuls composent peut-être la membrane et composent une membrane artificielle faite de lipides et protéines et en mesurent la tension superficielle. Ils observent que la valeur obtenue diminue pour se rapprocher de celle d’une vraie membrane plasmique. Conclusion : la bicouche lipidique n’est pas faite exclusivement de lipides mais aussi de protéines. Cela sera confirmé dans les années 1960 par des expériences de cryodécapage : on va décaper la bicouche lipidique et observer comme des petites granulations.
En fait, il existe des protéines insérées à l’intérieur de la membrane. Une protéine a une fonction. Sur le schéma bilan n’en a été représentée qu’une seule qui est une protéine canal qui laisse passer de manière discriminante certains ions ou certaines molécules.
IV. La membrane plasmique : une mosaïque fluide (Singer et Nicholson)
En 1972, Singer et Nicholson vont finaliser la composition de cette membrane : la membrane d’une cellule c’est une bicouche lipidique (des phosphoglycérolipides) dont les lipides qui la composent ont la capacité de bouger à la surface de leur couche, et parfois d’une couche à l’autre, on parle de flip flop. Enchâssées dans cette bicouche lipidique, il y a de nombreuses protéines.
Si on parle en pourcentage massique, elles composent 50 % de la membrane plasmique, alors que les lipides en composent 42 % et les glucides 8 %.
Cette membrane est en mouvement, ce qui permet aux protéines de bouger et permet la communication des cellules entre-elles. Nous ne sommes pas dans un modèle de cellule fermée avec l’extérieur : des molécules peuvent y pénétrer via des protéines-canaux, les protéines peuvent bouger au sein de la membrane. Cette membrane est en constant turn-over et en communication constante avec l’extérieur et intérieur de la cellule. Ce modèle est dit de la mosaïque fluide.