ATP et conversion d’énergie dans les cellules



ATP et conversion d’énergie dans les cellules
Il existe différentes formes d’énergie dans les cellules vivantes. Par exemple :
– L’énergie chimique : énergie contenue dans les molécules complexes chimiques, qui peut être libérée lorsqu’une molécule complexe est divisée en molécules plus petites.
– L’énergie calorique : énergie liée au dégagement de chaleur au cours de certaines réactions.
– L’énergie mécanique : énergie utilisée lors de mouvements cellulaires, par exemple le battement du flagelle des spermatozoïdes. Il peut s’agir également de l’énergie dissipée lors de la contraction musculaire.
– L’énergie osmotique : énergie liée à des différentiels de gradients de concentrations. Par exemple dans le cas d’un différentiel de concentration d’un ion entre un compartiment et un compartiment voisin, l’ion situé le compartiment où il est le plus concentré va avoir tendance à se déplacer vers le compartiment dans lequel il est le moins concentré. Ce déplacement spontané va éventuellement produire de l’énergie.
Une cellule vivante a donc besoin, en fonction du type cellulaire, de différents types d’énergie. Il existe dans nos cellules une molécule dite universelle, une sorte de monnaie d’échange énergétique : l’ATP.
ATP signifie adénosine triphosphate, il s’agit d’une molécule riche en énergie chimique.
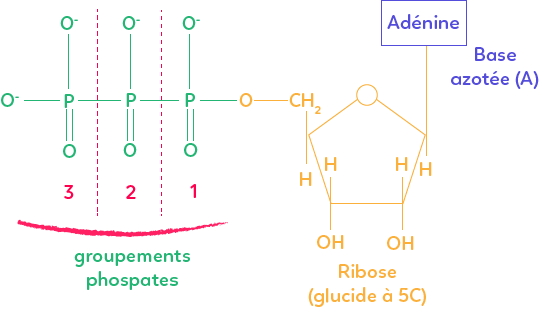
L’ATP est composée d’une base azotée, l’adénine, reliée à un sucre à 5 carbones, le ribose, lui-même relié à trois groupements phosphates.
L’énergie de cette molécule est principalement due à la force des liaisons covalentes entre les groupements phosphates. L’ATP libère de l’énergie sous forme chimique lorsqu’il est hydrolysé, c’est-à-dire lorsqu’il perd un ou deux groupements phosphates. C’est la rupture des liaisons covalentes entre les groupements phosphates qui libère de l’énergie rendant elle-même possible d’autres réactions consommatrices d’énergie.
Il existe donc en permanence des conversions d’énergies d’un type à un autre dans les cellules : l’ATP fait figure d’intermédiaire, il s’agit de la monnaie énergétique principale de nos cellules.
L’ATP peut être obtenue à partir d’une molécule semblable mais qui ne possède que deux phosphates, l’ADP (adénosine diphosphate), additionné à un groupement phosphate Pi appelé phosphate inorganique.
L’ADP peut être obtenu à partir d’une molécule semblable à l’ATP mais qui possède un seul groupement phosphate, l’AMP (adénosine monophosphate), additionné à un phosphate inorganique.
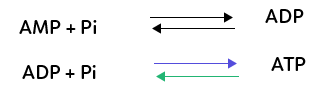
Il faut noter toutefois que la plupart des conversions dans nos cellules s’effectuent entre ADP et ATP.
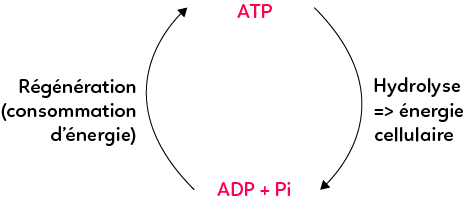
Le travail cellulaire correspond à une production permanente d’ATP qui rend possible les réactions consommatrices d‘énergie. L’ATP produit à partir d’ADP s’appelle une régénération. La régénération est consommatrice d’énergie. Elle est donc nécessairement couplée à d’autres réactions chimiques qui produisent de l’énergie, comme par exemple les oxydations des substrats organiques lors de la respiration cellulaire ou des fermentations, qui libèrent de l’énergie. À l’inverse, l’hydrolyse de l’ATP par la cellule, la séparant en ADP et Pi, libère l’énergie nécessaire aux réactions consommatrices d’énergie. Il s’agit donc d’un cycle entre la régénération permanente d’ATP et la consommation d’ATP par les cellules, qui permet aux autres réactions cellulaires d’avoir lieu.
ATP et contraction musculaire
Les muscles squelettiques sont attachés aux os de notre squelette, comme les muscles des bras ou les muscles des jambes, le quadriceps situé dans la cuisse par exemple.
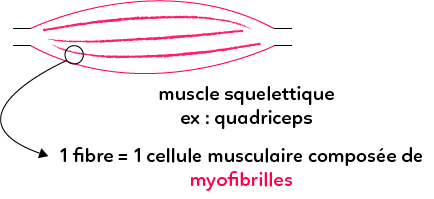
Ces muscles squelettiques sont constitués de très grandes cellules : les fibres musculaires. Elles peuvent atteindre jusqu’à 30 cm de long. Leur observation au microscope permet de repérer des myofibrilles à l’intérieur du cytoplasme des cellules musculaire, qui ont un aspect strié. On parle ainsi de muscles striés squelettiques et de cellules musculaires striées squelettiques.
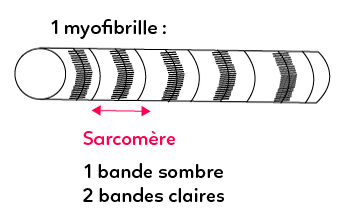
Cet aspect strié est dû à une répartition de filaments particuliers appelés les myofilaments, internes à la cellule musculaire striée squelettique. Cette organisation, caractéristique de la cellule musculaire, repose sur la combinaison de filaments épais (filaments de myosine) croisés avec des filaments fins filaments d’actine). Chacune des unités répétitives observées le long de la myofibrille sont les sarcomères. Le sarcomère sur le schéma ci-dessus est encadrés par deux stries (constituées elles aussi de molécules protéiques) appelées les stries Z.
État relâché et état contracté
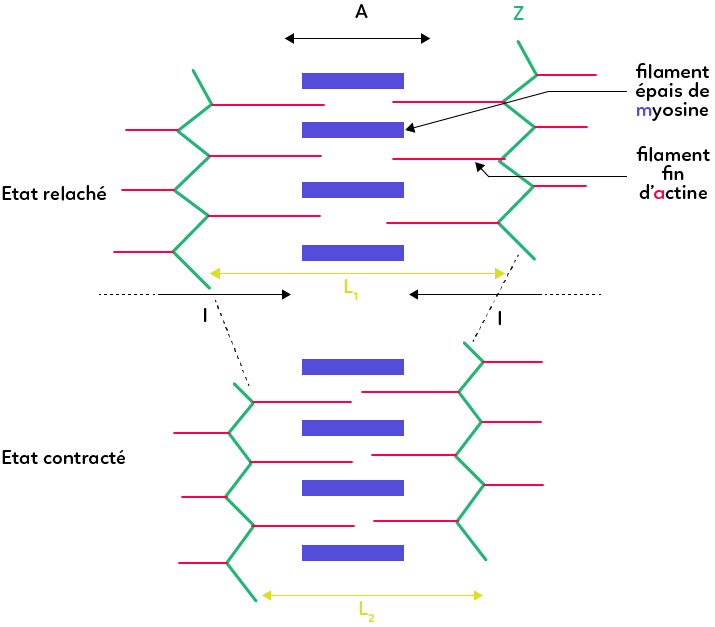
Le cœur du sarcomère en microscopie est plus sombre, c’est ce qu’on appelle la bande A.
A l’état relâché, la bande A correspond à une superposition de myosine (en bleu dans le schéma) et d’actine (en rouge).
On observe en microscopie des zones plus claires vers les extrémités des sarcomères : il s’agit de zones dénuées de superposition de myosine et d’actine mais composées uniquement des filaments fins d’actine, correspondant à ce qu’on appelle bande I ou disque I. Celle-ci est à cheval sur un sarcomère et son voisin, puisqu’il faut imaginer à gauche et à droite du schéma des successions d’autres sarcomères.
A l’état relâché, la longueur d’un sarcomère est d’environ 2 µm. Il est capable de se contracter. Lors d’une contraction musculaire, on observe un coulissage des myofilaments les uns par rapport aux autres et donc un rapprochement des deux stries Z, délimitant le sarcomère. Cependant, la longueur du filament de myosine (longueur A) ne varie pas. C’est la longueur des disques I qui diminue au fur et à mesure que les myofilaments coulissent les uns par rapport aux autres. Par conséquent, les sarcomères les uns à la suite des autres se rapprochent mutuellement : on obtient au final un myofilament plus court et donc un muscle long à l’état relâché et un muscle court à l’état contracté.
Mécanismes à l’œuvre dans cette contraction musculaire
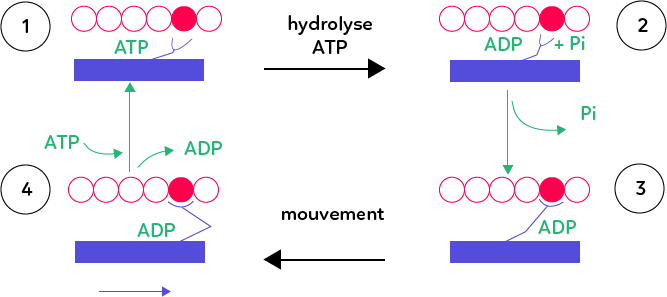
Les filaments fins d’actine sont représentés en rouge et les filaments de myosine en bleu. Au cours de la contraction, les myofilaments fins et épais s’attachent temporairement l’un à l’autre. Cet attachement, qui permet ensuite le coulissage, requiert la présence d’ATP. En effet, l’ATP est hydrolysée juste avant que les filaments ne bougent l’un par rapport à l’autre.
Au départ (étape 1) la molécule de myosine est liée par ce qu’on appelle les têtes de myosine a une molécule d’ATP. Celle-ci est ensuite hydrolysée (découpée) en de l’ADP et du phosphate inorganique (Pi) par les têtes de myosine (étape 2). Ce découpage permet d’activer la molécule de myosine qui libère son phosphate inorganique dans un second temps.
La molécule de myosine prend alors une conformation qui lui permet de s’attacher temporairement à un endroit précis du myofilament fin d’actine (étape 3). Les deux filaments sont alors liés l’un à l’autre. La myosine reste par ailleurs liée à de l’ADP.
Cet attachement des deux filaments entre eux débouche sur un mouvement, un coulissage (étape 4). Sur le schéma, le myofilament fin se déplace légèrement vers la gauche et le myofilament épais vers la droite. Ce coulissage est rendu possible par la bascule de la tête de myosine, dont son angle par rapport à l’actine est modifié. Par des mouvements successifs, identiques à celui décrit, on obtient alors un rétrécissement progressif du sarcomère.
Dans la dernière étape, la molécule d’ADP est libérée ce qui déclenche le décrochage du myofilament épais et du myofilament fin. L’ATP vient reprendre la place de l’ADP au sein de la molécule de myosine.
On revient alors à l’étape 1 du schéma : un nouveau cycle peut avoir lieu, où la tête de myosine vient s’attacher un peu plus loin sur la molécule d’actine. De proche en proche, en s’accrochant successivement plus loin sur la molécule d’actine, la myosine coulisse alors progressivement et raccourcit la longueur du sarcomère.
Forte consommation d’ATP
On remarque ainsi que tout le cycle nécessite une forte consommation d’ATP. Tout l’enjeu pour la cellule musculaire est de régénérer en permanence l’ATP. Une cellule ne dispose pas stock d’ATP et a besoin au fur et à mesure de son effort de reconstituer l’ATP.
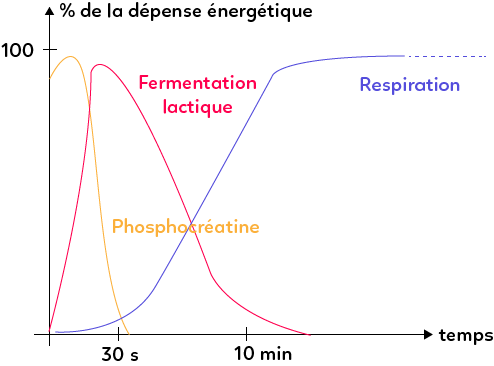
Pour les efforts musculaires de quelques secondes, qui sont les plus violents (efforts d’haltérophile par exemple) l’ATP est régénérée du stock de phosphocréatine qui fournit son phosphate, combiné à l’ADP pour former de l’ATP.
Si l’effort est un peu plus long, quelques dizaines de secondes voire quelques minutes, l’ATP est régénérée par fermentation lactique. Les cellules musculaires sont capables de fermenter, mais elles produisent de l’acide lactique responsable de la fatigue musculaire, comme les crampes. Il s’agit donc d’un mécanisme qui est capable de régénérer un peu d’ATP mais qui n’est pas durable dans le temps.
Le troisième mécanisme, le plus efficace à long terme, est celui de la respiration cellulaire. Lors de celle-ci, la cellule réalise la production de 36 molécules d’ATP par glucose oxydé. Cela permet alors d’avoir une régénération plus longue de l’ATP et de fournir un effort musculaire, donc une contraction des sarcomères, de façon plus durable.